Electron transport chain
2.79M views702 WordsCopy TextShare
Harvard Online
From our free online course, “Cell Biology: Mitochondria”: https://www.edx.org/course/cell-biology-m...
Video Transcript:
RL: All eukaryotic cells, from yeast to those that make up the human body, contain membrane-bound organelles with specialized functions. Mitochondria are double-membraned organelles that harness most of the energy that cells need to grow and reproduce. Nearly all of this energy comes from reactions that take place at the inner mitochondrial membrane.
One of the key roles of this membrane is to act as a barrier to positively charged particles called protons, thus allowing a concentration gradient to be maintained where the intermembrane space has far more protons than the matrix. The membrane also contains a large protein complex called the F1F0 ATP synthase, which uses the proton gradient to drive the synthesis of ATP molecules. These ATP molecules ultimately provide the energy for most of the cell's reactions.
Just as man-made power plants produce electrical energy by using the flow of wind, water, or steam to rotate a turbine, the synthase makes ATP by using proton flow from one side of the inner membrane to the other to rotate protein subunits. If there is no proton gradient, synthase subunits stop rotating, and the cell can quickly become starved of the energy and die. Therefore, the protein complexes and small molecules that establish this gradient and maintain it play an essential role in the life of the cell.
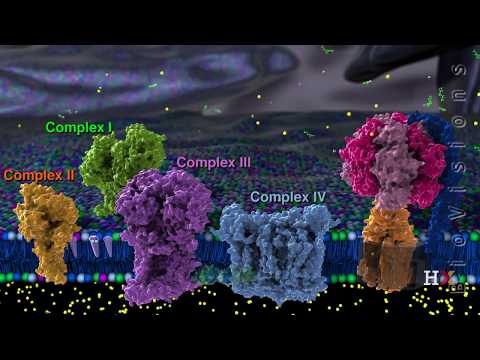
At the heart of this system are four protein complexes numbered I through IV. Complexes I, III, and IV directly pump protons from the matrix into the intermembrane space. Complex II does not directly pump protons, but it does promote proton pumping in complexes III and IV.
Proton pumping requires energy, and the four protein complexes get this energy by transferring electrons through a series of coupled reactions. This linked process of electron transport is why the four complexes are collectively referred to as the electron transport chain. Let's focus on complex I.
A byproduct of sugar metabolism called NADH deposits two high-energy electrons in complex I, where they are passed along a chain of redox centers. Redox centers are clusters of atoms that have different affinities for electrons based on their unique atomic configurations. Let's closely consider a pair of redox centers to reveal two reasons why an electron moves from the top redox center to the bottom.
First, the bottom redox center has higher affinity than the top one. Second, the distance between these adjacent redox centers is ideal for an electron jump to occur, which explains why electrons typically don't bypass the bottom redox center. A small amount of energy is released each time an electron is passed between redox centers.
Complex I harnesses this energy across all the redox centers and uses it to pump protons. The last redox center in complex I donates two electrons to a coenzyme Q molecule. Complex II is similar to complex I in two important ways.
First, high-energy electrons also enter complex II via a byproduct of sugar metabolism, although here the molecule is FADH2. Second, complex II also transfers electrons between several redox centers before donating them to coenzyme Q. One major difference, however, is that complex II does not use the energy liberated to pump protons.
Coenzyme Q molecules from complexes I and II donate their electrons to complex III. One electron is a recyclable and can re-enter complex III later, but the other passes through two redox centers before reaching cytochrome c. Cytochrome c carries the electron to complex IV.
The electron transport chain ends in complex IV, where a series of reactions involving four electrons converts a molecule of oxygen to two molecules of water. The proton gradient is strengthened because four protons from the matrix are incorporated into water molecules, and another four are pumped into the intermembrane space. In the absence of oxygen, the electron transfer comes to a halt, meaning that ATP synthesis also stops.
Indeed, the reason we breathe oxygen is so that it can serve as the final electron acceptor at the end of the electron transport chain. In this animation, we have explored each protein complex in isolation, but in reality, they are very densely packed. Together, they effectively make the entire surface of the inner mitochondrial membrane a giant cellular power plant.
Related Videos
4:59
ATP synthase in action
Harvard Online
1,406,028 views

16:52
Electron Transport Chain (Oxidative Phosph...
Dirty Medicine
1,916,845 views

5:44
Mitochondria: the cell's powerhouse
Harvard Online
1,051,896 views

7:20
DNA animation (2002-2014) by Drew Berry an...
WEHImovies
5,702,356 views
7:30
Immunology in the skin
nature video
2,936,829 views

7:45
Electron Transport Chain
Omar Ali
1,417,564 views

5:02
How a synapse works
Harvard Online
1,384,124 views
4:53
The Electron Transport Chain Explained (Ae...
BioMan Biology
741,154 views

6:58
What is a Protein? (from PDB-101)
RCSBProteinDataBank
2,864,364 views
9:09
Animations of unseeable biology | Drew Ber...
TED
2,558,256 views

16:41
The Insane Biology of: Slime Mold
Real Science
1,050,331 views
14:02
Cellular Respiration: Do Cells Breathe?: C...
CrashCourse
120,588 views

18:52
The Science Of Self Control
HealthyGamerGG
1,251,225 views

29:22
Metabolism | Electron Transport Chain: DET...
Ninja Nerd
737,244 views

9:44
Mitochondria Aren't Just the Powerhouse of...
Seeker
504,027 views

20:47
The Genius Behind the Quantum Navigation B...
Dr Ben Miles
999,882 views

11:31
Electron Transport Chain | Made Easy
Dr Matt & Dr Mike
143,659 views
6:01
Mysteries of the Electron Transport Chain,...
sciencemusicvideos
965,259 views

8:47
Cellular Respiration (UPDATED)
Amoeba Sisters
4,116,711 views

7:37
The 5 core principles of life | Nobel Priz...
Big Think
2,273,074 views